
Alimentation et dépendance
Le cerveau est le centre du système nerveux. Il est capable d’intégrer les informations, de contrôler la motricité et d’assurer les fonctions cognitives.
Les centres du circuit de la récompense agissent ensemble afin de guider nos actions et notre comportement. On observe ainsi des interactions entre les différentes aires cérébrales impliquées dans le circuit.
Par quel moyen ces différentes régions du cerveau communiquent ?
Cette communication se fait par le biais de cellules nerveuses, appelées neurones. Les neurones établissent des connections, des liens entre eux afin de créer de véritables voies d’informations contrôlant différents aspect de notre vie (apprentissages, mouvements, émotions…).
Les cellules nerveuses sont constituées de dendrites (correspondant à la voie d’entrée du message nerveux) et d’un axone (voie de sortie), qui peut être recouvert par une gaine de myéline (enveloppe isolante faite d’une substance riche en lipides (la myéline) permettant à l’influx nerveux de circuler plus rapidement).
Entre, on trouve l’élément central de la cellule : le corps cellulaire (cell body), aussi appelé le soma. Le soma correspond à la partie sphérique du neurone et contient le nucléus (noyau de la cellule).
Lorsque l’information d’un autre neurone est reçue, les dendrites transmettent le signal au soma qui par la suite en fonction de la puissance de ce signal, le transmet à l’axone.
Le cerveau est un mélange de neurones :
- autoactifs qui produisent des décharges électriques par bouffées de potentiels d'action (Bursting neurons)
- produisant des décharges à fréquence constante (tonic firing)
- silencieux (mais pouvant être stimulés)
Bien d’autres critères permettent de différencier certains types de neurones tels que leur quantité de production de potentiel d’action par exemple.
Le fonctionnement cérébral ou les apprentissages changent:
- la force des synapses ou la création de nouvelles synapses.
- le mode ou la fréquence de décharge des neurones.
Toutes ces modifications peuvent se produire sur des neurones différents ou sur le même neurone.
La communication entre deux neurones est assurée par des jonctions particulières, les synapses correspondant à un espace entre deux neurones appelée fente synaptique. La largeur de ces fentes est inférieure à 0.1 micron (100 millionièmes de mètre) et le temps de passage du message d’une cellule à l’autre est de l’ordre de 0.5 millisecondes.
Les synapses peuvent être de plusieurs types :
-Chimique (B) dont le message est transmis d’un neurone à l’autre par l’intermédiaire de neurotransmetteurs.
-Electrique (A), dont le message est transmis par l’intermédiaire de jonctions communicantes de type GAP reliant les éléments pré et post-synaptique. La seule différence anatomique entre ces deux types de synapses est due à la jonction GAP de la synapse électrique.
- Excitatrices ou inhibitrices.
- Vers des muscles (synapses neuro-musculaires) ou vers d’autres neurones (synapses neuro-neuronales) .
Ici, on s’intéresse aux synapses chimiques neuro-neuronales dans le contexte du circuit de la récompense.
Le neurone d’où arrive l’influx nerveux est appelé pré-synaptique. Celui où vont se fixer les neurotransmetteurs porte le nom d’élément post-synaptique.
Comment se forment les synapses ?
Les cellules sont constamment à la recherche d’un contact permanent et stable. Ce contact ne peut se produire que lorsque deux cellules sont maintenues en place par des molécules d'adhésion.
De plus, il est important de maintenir un espace très restreint entre ces deux cellules (environ 0,1 microns). Dans le cas contraire, la transmission des signaux neuronaux serait inefficace (due à la dilution des neurotransmetteurs et au temps de diffusion de ces derniers relativement long).
Un certain nombre de protéines transmembranaires jouent le rôle des molécules d'adhésion.
Chez les neurones, une famille de protéines neuronales, les neurexines, en plus de jouer un rôle dans le fonctionnement des synapses (synaptogénèse / maturation synaptique) interviennent dans le mécanisme d’adhésion.
Une autre protéine nommée SynCAM (protéine située de chacun des 2 côtés des neurones impliqués) possédant une extrémité intracellulaire homologue de celle des neurexines et une partie extracellulaire très caractéristique qui permet l'initiation de la jonction. Les autres protéines (présentes sur le schéma) permettent de renforcer, maintenir cette adhésion.
Nous avons représenté ces complexes sur les schémas ci dessous.
LA SYNAPSE

Schéma d'un réseau neuronal

Modification des connexions noeurnales suite à un apprentissage

Schéma présentant les différences entre les deux types de synapses (électrique et chimique)


Complexe neurogiline/β-Neurixine avec les N et C terminaux apparents
Après que les molécules d'adhésion aient préparé la structure, les récepteurs, les vésicules et la machinerie de fusion des vésicules doivent être recrutés par les synapses . Sinon, la synapse ne fonctionnera pas.
Lorsque les synapses sont très sollicitées, celles-ci deviennent plus grosses et possèdent plus de récepteurs et de vésicules de sécrétion, de sorte à ce que le changement d'une transmission productive des actions augmente la transmission du potentiel d'action (synapse stimulatrice) ou le diminue dans le cas contraire (synapse inhibitrice).
Comment se transmet le message?
1/ le Potentiel d'Action (PA) :
Le potentiel d’action est constitué de trois phases principales. Premièrement la dépolarisation, d’une amplitude de 100mV faisant passer le potentiel de la membrane de -70 à +30mV. En suit une repolarisation qui correspond à un retour au potentiel de repos.
En général, le potentiel atteint une valeur plus basse que celle de référence -80mV, c’est l’hyperpolarisation diminuant la fréquence du potentiel d’action (on parle de période réfractaire) pour ensuite retourner à la valeur standard, -70mV. Cette hyperpolarisation est la conséquence des canaux Na+ voltage dépendant fermé pendant une période très brève.
Le long d’un neurone, l’influx nerveux est transmis sous la forme d’une séquence de potentiel d’action. Le message est donc en premier lieu de nature électrique.
Le potentiel d’action est un signal électrique unidirectionnel traversant le neurone jusqu’à l’axone, événement durant lequel la polarité de la membrane s'inverse. Ce signal électrique dure entre 1 et 2 millisecondes.
Le potentiel transmembranaire au repos appelé potentiel de repos a une valeur d’environ -70mV.
Le potentiel d'action est constitué de trois phases principales. Premièrement la dépolarisation, d’une amplitude de 100mV faisant passer le potentiel de la membrane de -70 à +30mV. En suit une repolarisation qui correspond à un retour au potentiel de repos.
En général, le potentiel atteint une valeur plus basse que celle de référence -80mV, c’est l’hyperpolarisation diminuant la fréquence du potentiel d’action (on parle de période réfractaire) pour ensuite retourner à la valeur standard, -70mV. Cette hyperpolarisation est la conséquence des canaux Na+ voltage dépendant fermé pendant une période très brève.
C’est au niveau du cône axonique (ou cône d’émergence de l’axone), que ce forme le potentiel d'action. Lieu où les canaux potassiques et sodiques sont en surabondance au niveau de la membrane plasmique. Lorsque les dépolarisations somato-dendritiques y arrivent et que leur sommation atteignent ou dépassent le potentiel seuil, il se produit une ouverture de tous canaux (seuls les canaux qui sont adjacents à la zone de dépolarisation ou ceux qui sont dans le « nœud de Renvier » (espace entre deux gaines de myéline) pour les neurones myélinisés) sodiques voltage-dépendant donnant naissance à des potentiels d'action qui vont se déplacer le long de l'axone.
Le potentiel d'action parvient ensuite jusqu'à la pointe de l'axone afin d'activer, au niveau de la synapse les canaux Ca2+ voltage dépendant, affectant la sécrétion de neurotransmetteurs.
Ce potentiel d'action est un phénomène électrique qui présente deux caractéristiques fondamentales :
- A partir de la valeur seuil de dépolarisation, toute augmentation ultérieure de l'amplitude de la stimulation n'apporte aucun changement dans la réponse observée : le PA obéit à la loi du tout ou rien. Si le seuil de dépolarisation n'est pas atteint, il n'apparaît pas. Si le seuil est atteint, la réponse est maximale d'emblée. Emis en un point de l'axone, il se propage sans atténuation tout au long de la fibre.
- Le déplacement d'un potentiel d'action le long d'un axone, modifie succéssivement la polarité de la membrane cellulaire.


Animation du déplacement du potentiel d'action le long d'un axone
Représentation d'un potentiel d'action

Étapes détaillées du déplacement du potentiel d’action :
a) dépolarisation
L’arrivée d’un stimulus engendre l’ouverture d’un nombre croissant de canaux voltage-dépendants à Na+ laissant entrer des cations dans le milieu intracellulaire. Les charges positives se regroupent du côté interne de la membrane. Ceci provoque une légère dépolarisation de la membrane
Lorsque la dépolarisation est suffisante pour atteindre le potentiel seuil, tous les canaux voltage-dépendants à Na+ sont ouverts provoquant une entrée brutale des ions Na+ dans la cellule nerveuse. Ceci provoque un retournement du potentiel de la membrane à une valeur de +30mV.
b) repolarisation
La forte dépolarisation finit par désactiver les canaux sodiques les refermant ainsi (période réfractaire). Les canaux K+ voltage-dépendants s’ouvrent lorsque le voltage de la membrane dépasse les +20mV. Quand le flux entrant de Na+ est arrêté, le flux des ions K+ sortant va remettre la polarité vers une valeur négative.
c) hyperpolarisation
Le fort flux sortant très important de K+ peut occasionner une hyperpolarisation transitoire de la membrane (valeur de -80mV), pendant cette période il est plus difficile de déclencher un nouveau potentiel d’action.
d) retour au potentiel de membrane
Les canaux K+ voltage-dépendants se ferment progressivement jusqu’à atteindre le nombre initial de canaux ouverts au repos. La membrane récupère ainsi son potentiel de repos. Les concentrations ioniques initiales sont restaurées dans un laps de temps supérieur à la durée du potentiel d’action grâce une pompe ionique sodium-potassium ATP dépendante.

2/ Le message chimique :
2.1 Vidéo
La synapse est un élément majeur du système nerveux et l'actrice principale des connections entre neurones. La vidéo qui suit vous explique son fonctionnenment plus en détails.
Pompe Na-K ATPase :
Cette pompe, formée de deux sous-unités assemblées en un hétérodimère, transporte les ions Na+ et K+ dans le sens inverse de leur gradient électrochimique (K+ sort et Na+ entre) et ce grâce à l'énergie fournie par l'hydrolyse de l'ATP préalablement phosphorylé.
Draw my Synapse : Comment le potentiel d'action provoque la libération des neurotransmetteurs ?
2.2 Mécanisme permettant le passage d'un message électrique en un message chimique
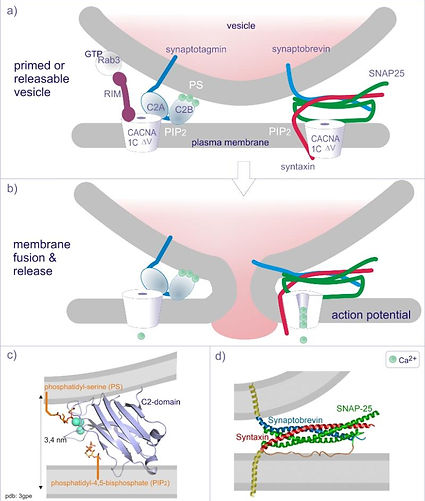
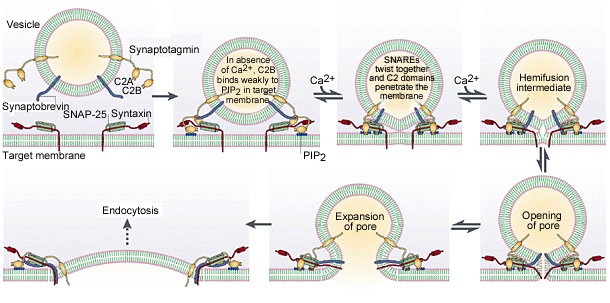
L'exocytose (fusion de la vésicule avec la membrane du neurone présynaptique permettant la libération des neurotransmetteurs dans la fente synaptique) est assurée par le complexe SNARE :
L’arrivée de calcium permet l’enroulement de la synaptopbrévine (associée à la membrane de la vésicule) avec les protéines SNAP 25 et syntaxine (associées à la membrane plasmique). Cela permet un arrimage de la vésicule à la membrane de la cellule. Ca2+ se fixe alors à la synaptotagmine, présente dans la membrane de la vésicule renforçant l’interaction entre la vésicule et la membrane plasmique. Ensuite, le flux entrant de Ca2+, par le canal CANCNA1C provoqué par l’arrivé de potentiel d’action induit la fusion des deux membranes.
1/
2/
Schématisations du fonctionnement du complexe SNARE
Remarques:
-La compréhension du rôle de Ca2+ dans la fusion membranaire n'est pas totale (des doutes sur le modèle actuel persistent).
-Le domaine C2B dispose de deux sites de liaison: l'une est pour PIP2 (phosphatidylinositol-3,4-bisphosphate) un inositol lipide présent dans la membrane plasmique, l'autre est avec la phosphatidylsérine (PS), mais cela ne se produit qu'en présence de Ca2+. En d'autres termes, Ca2 + provoque une fixation ferme de la membrane de la vésicule à la membrane plasmique. Ensuite, la fusion des deux membranes nécessite un changement dans les protéines du complexe SNARE (SNAP-25, Synaptobrevin et syntaxine) .
-La réflexion actuelle est que l'ouverture du voltage canal de Ca2+ (CACNA1CDV, voir schéma 1) et le passage ultérieur des ions Ca2+ dans le canal provoque un changement dans le «complexe de fusion» qui provoque la fusion membranaire (la position du segment transmembranaire de syntaxine, en rouge change, voir schéma 1).
2.3 Mécanisme permettant le passage d'un message chimique en message électrique
-Récepteurs ionotrope : voir vidéo
-Récepteurs métabotropes : de manière indirecte suite à l'activation de la protéine G.

Expliquation schématique des récepteurs métabotropes cités dans la vidéo ci dessus
Ces récepteurs sont à l'origine d'une réponse plus lente et prolongée (allant de quelques millisecondes à quelques minutes). Les différentes cascades de réaction présentées dans le schéma ci dessus expliquent ce délai d'action plus long. Certaines de ces réactions conduisent à l'activation de protéines régulatrices issues de la transcription génique (la durées d'action peut ainsi s'étendre sur plusieurs mois). Les neurotransmetteurs participent alors au codage à long terme des informations nerveuses (détaillé dans le paragraphe ci dessous). Les effets postsynaptiques liés aux récepteurs couplés aux protéines G sont donc plus lents, plus durables et plus variés que les effets liés aux récepteurs ionotropes.
La potentialisation à long terme - synapse et mémoire :
Et si nous vous disions que les synapses sont effectivement utiles pour notre mémoire ? Celles ci effectuent en effet une potentialisation à long terme (PTL). Après une intense activation pré-synaptique (fréquence supérieure à 100Hz en moins d'une seconde), une réaction en chaine se met en place, initiée par une entrée de calcium dans l'axone du neurone pré-synaptique (se référer à la vidéo). La PTL qui s'amorce alors comporte deux phases. L'établissement, ou induction induite par la forte fréquence de l'influx nerveux dure environ une heure et implique l'action de plusiqueurs enzymes qui persisteront après l'élimination du calcium (Ca2+ ). Une phase de maintien s'en suit, pouvant durer plusieurs jours, mise en place après une succession de stimulations. Elle conduit à la synthèse de protéines. Des "souvenirs" peuvent alors être encodés à l'intérieur du réseau de neurones par les synapses, et essentiellement les synapses chimiques qui sont plus souples et malléables, caractéristiques précieuses à la base de tout apprentissage.
Sources :
Site traitant de l'anatomie et de la physiologie du SNC
hhmi Howard Hugues Medical Institute
http://www.afblum.be/bioafb/neurone/neurone.htm
http://www.museum-marseille.org/marseille_cerveau_synapse.htm
http://neurobranches.chez-alice.fr/neurophy/lasynapse.html
http://neurobranches.chez-alice.fr/neurophy/potact.html
https://fr.wikipedia.org/wiki/Synapse
https://sites.google.com/site/aphysionado/home/cellsnv/PA/recptrneurotransm
http://lydia.danglot.free.fr/Cours/Master2-P6-Danglot.pdf
http://www.pourlascience.fr/ewb_pages/a/article-un-cerveau-jamais-au-repos-25342.php
http://www.larecherche.fr/actualite/vie/proteine-a-origine-synapses-01-11-2002-82457

